
Une aventure scientifique et humaine

Une aventure scientifique autour de la biodiversité : de la forêt amazonienne à l’évolution, en passant par la modélisation et la gestion des écosystèmes. L’aventure amazonienne avec le CNRS.
Rev. octobre 2011
Aventures
Septembre 1993 à la station des “Nouragues” en Guyane française, assis sur un petit banc au bout de la DZ, ou « Dropping Zone », aire d’atterrissage des hélicoptères qui assurent la liaison avec Cayenne, je contemple un minuscule morceau de la forêt Guyanaise, elle-même ne représentant qu’une faible part du grand système forestier amazonien. C’est un peu avant la tombée de la nuit et j’espère voir quelques animaux, notamment des singes. Ceux-ci se font attendre et une fois de plus, au soleil couchant, j’admire la symphonie de verts, qui traduit une part de la diversité des arbres de cette forêt. Même sans être botaniste, la vue des couleurs, combinée à celle de la forme des houppiers indique que devant moi il y a beaucoup d’arbres d’espèces, ou essences pour prendre le langage des forestiers, très différentes et plus encore que des arbres voisins semblent appartenir à des catégories distinctes. Je l’avais déjà vu lors de missions précédentes, mais n’étant pas spécialiste et même si j’étais étonné de cette diversité, je la prenais comme un fait, fruit du hasard des choses ou de processus qui m’étaient étrangers et sans doute bien connus de mes amis écologues.
Les animaux se faisant toujours attendre, je me demandais ce qui pouvait expliquer cet apparent désordre arboricole. Faisant métier de modélisateur, métier à l’époque dont on commençait tout juste à cerner les contours, la traduction en était : « quel modèle (simple !) pourrait représenter cette structure spatiale ? ». Bien qu’ayant, en tant qu’enseignant et que chercheur, eu à faire avec le hasard du calcul des probabilités et de la statistique, j’avoue avoir été un ardent partisan des modèles déterministes et j’en ai commis quelques-uns qui se sont avérés utiles. Mais revenons aux arbres des Nouragues, tout en attendant ces primates qui ne venaient décidément pas, je commençais à rêver… Peut-être que du chaos (déterministe bien sûr !) pourrait rendre compte de cette structure désordonnée. Peut-être après tout que le modèle de base ne serait pas très compliqué. En effet, un modèle aussi simple que le modèle logistique en temps discret peut engendrer une dynamique d’apparence erratique. Puis vint l’analogie musicale, certaines œuvres paraissent compliquées et harmonieuses alors que le « motif de base » est simple. Pour un canon, il suffit d’une répétition après un décalage, par exemple d’une mesure. Souvenons nous du « Frère Jacques » de notre enfance ou de « l’Offrande Musicale » de Jean-Sébastien Bach. C’était le début d’une nouvelle aventure scientifique et il a fallu une dizaine d’année pour que je puisse avancer une explication raisonnable. C’est vrai aussi qu’au départ, il s’agissait pour moi plus d’une curiosité que d’une préoccupation scientifique. J’avais d’autres choses à faire et plus urgentes, notamment de préparer ce qui devait devenir le « Programme Environnement, Vie et Sociétés » du CNRS et visser quelques écrous pour finaliser un livre sur la modélisation des systèmes biologiques et écologiques, fruit des vingt premières années de mon activité scientifique et pédagogique [1].
Nous sommes maintenant en 1996. Je suis à nouveau dans l’avion en direction de la Guyane, je feuillette un des derniers numéros de Nature et je tombe en arrêt devant un article signé par Vincent Courtillot et Yves Gaudemer sur la modélisation de l’évolution de la biodiversité depuis 600 millions d’années [2]. Le modèle choisi est une vieille connaissance : le modèle logistique, mais là en temps continu. J’ai toujours été émerveillé par le fait que ce modèle simple représente une grande diversité de situations biologiques, remarque déjà faite par Feller dans les années 1940. Plus récemment j’avais montré qu’il pouvait représenter plusieurs types de grands mécanismes biologiques et écologiques. C’était aussi un très bon outil pédagogique me servant d’introduction à la modélisation pour les étudiants biologistes : mathématiquement il ne présente pas de difficultés majeures ; en revanche, on peut introduire des concepts de base des « systèmes dynamiques » chers aux mathématiciens et aux physiciens. “Super ! me suis-je dit, un nouvel exemple à présenter à mes étudiants et là, ce ne sont plus des dynamiques de populations microbiennes dans un tube à essais mais quelque chose à grande échelle d’espace et de temps : la planète et les ères géologiques”.
Et puis… au lieu d’en rester là. J’ai lu l’article avec un intérêt grandissant en notant mon accord sur l’essentiel, mais aussi qu’il était possible d’apporter quelques améliorations notamment sur le plan méthodologique et peut-être aussi d’utiliser ce que j’avais démontré sur l’utilisation du modèle. J’aurai pu me contenter de ces remarques, mais non. Après ma mission, j’ai écrit une lettre à Vincent Courtillot qui me répondit pratiquement par retour du courrier en m’encourageant à poursuivre. Deux collègues m’ont accompagné dans une partie de cette aventure : Jean-Christophe Hervé, un de mes anciens doctorants, excellent spécialiste des modèles non linéaires, et Claudine Schmidt-Lainé, une collègue, consœur et amie mathématicienne, qui s’illustre dans les recherches sur l’environnement.
Par ailleurs, j’avais entendu parler d’évolution et lu quelques articles et ouvrages convaincants sur la question, mais pour apporter une contribution significative au débat, tout en étant fort occupé par ailleurs, il a fallu 6 ans pour publier l’article présentant les résultats (2002) [3].
Quatre ans de plus ont été nécessaires pour que la relation entre ces résultats sur la dynamique de la diversité biologique et ce que j’avais observé sur la biodiversité et l’hétérogénéité des forêts tropicales puisse se concrétiser. Cependant, il est bon de faire une remarque : dans mes fonctions d’animation et de gestion de la recherche, j’ai été confronté aux deux grandes problématiques actuelles de l’environnement global, celles du climat et celle de la biodiversité. La façon d’aborder ces problématiques par la communauté scientifique est assez différente. Pour le climat, la construction d’un discours et de modèles globaux a été le souci principal. On sort des particularismes locaux, quitte à y revenir par la suite, mais comme instance locale d’un modèle général. En revanche, pour la biodiversité et malgré des débuts prometteurs, on peine à élaborer une vision globale à la fois dans l’espace, du local au planétaire, entre les niveaux d’organisation biologique, « du gène à l’écosystème », et dans le temps, celui d’une génération à celui de l’évolution des êtres vivants. Mon souci principal est qu’un tel discours soit ébauché pour jeter les bases d’une théorie synthétique de la biodiversité, notamment en soutenant ceux et celles qui veulent y contribuer. Pour ma part, sans que j’en aie eu l’intention affirmée, j’ai tenté d’y apporter ma pierre en réalisant un ouvrage [4], complété et traduit en anglais [10] et un article [5]. Et sans que ce fût l’objectif, cette contribution n’est pas sans retombées sur la théorie de l’évolution. Bien sûr le discours intègre le fait évolutif, mais sans que ce soit le centre du discours et ce qui m’a attiré l’attention, c’est qu’une des premières critiques de mon ouvrage, au demeurant très positive, est parue dans un numéro des « Dossiers de La Recherche » consacré à l’Évolution [6].
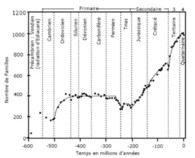
Figure 1 - Évolution de la biodiversité marine au cours des 600 derniers millions d’années. Les données ont été extraites de la base de données constituée par un collègue américain J.J. Sepkoski et utilisées par Y. Gaudemer et V. Courtillot dans leur article. La courbe en continu correspond à une modèle logistique par morceaux : l’expression analytique est la même, ce sont les valeurs des paramètres qui diffèrent. Dans l’article de Y. Gaudemer et de V. Courtillot, seules les phases croissantes étaient modélisées. Notre contribution a porté sur l’estimation des paramètres et la précision de ces estimations, sur la modélisation simultanées des phases croissantes et décroissantes et sur l’interprétation mécaniste du modèle. On notera enfin, à l’échelle géologique et en tendance moyenne, que la biodiversité croit de façon quasi exponentielle, malgré les « grandes extinctions
Revenons à la biodiversité. Son histoire, telle que nous pouvons la reconstituer, montre qu’en moyenne elle est en croissance continue et quasi exponentielle (figure 3). Cette histoire nous montre aussi que les êtres vivants deviennent, toujours en moyenne, plus complexes. Enfin, malgré de grandes catastrophes, qui se sont traduites par des diminutions drastiques de la diversité biologique, le processus de diversification a permis d’amortir ces chocs, de combler les diminutions et même d’augmenter la biodiversité. Enfin, quand on analyse les processus en cause, on voit qu’ils sont nombreux et surtout que le hasard y joue un rôle essentiel.
Concrètement, on se rend vite compte que la biodiversité est une assurance pour la vie. En effet, à un moment donné, avoir des individus très diversifiés augmente les chances qu’il en existe, a priori, pouvant survivre à des changements de l’environnement, même sévères. Ces survivants seront ensuite à l’origine de nouvelles diversifications, grâce au « hasard » qui est à la base des processus de diversification.
Longtemps considéré comme contingent et exogène aux systèmes biologiques, le hasard, dont nous parlons, a en fait deux origines, l’une externe et l’autre interne. Externe, d’origine environnementale, il se traduit par des événements à allure aléatoire qui modifient des systèmes biologiques, par exemple des mutations dues aux radiations ionisantes naturelles. Interne, il est produit pas des processus biologiques à tous les niveaux d’organisation du vivant. Ils sont d’origine biophysique, biochimique ou comportementaux. C’est par exemple le cas des migrations des chromosomes lors des divisions cellulaires : les nouveaux et anciens chromosomes se répartissent au hasard entre les cellules filles. C’est encore le cas lors d’échanges entre chromosomes homologues, par exemple par « crossing over ». C’est toujours le cas lors du choix des partenaires lors de la reproduction sexuée. Dans un autres registre, on observe la trajectoire erratique de proie fuyant devant un prédateur, comme celui de la gazelle devant un guépard. Enfin, les processus de dissémination des graines végétales ont une forte composante aléatoire, par exemple lorsque cette dissémination est assurée par les animaux. Cela assure une grande hétérogénéité spatiale dans la distribution des individus. La figure 4 illustre l’intérêt d’un système où les individus d’espèces différentes sont très mélangés. Ainsi et en résumé, le hasard est essentiel pour augmenter la biodiversité et la maintenir. La biodiversité est un facteur de survie pour les communautés d’êtres vivants. La trajectoire erratique d’une proie lui donne oplus de chances de survie que si elle allait droit devant elle. Le hasard est donc nécessaire à la vie et à la survie.
Les processus endogènes qui engendrent ce hasard, que nous avons appelés des « roulettes biologiques », sont apparus spontanément et ont été sélectionnés lors de l’évolution des êtres vivants, car il munissent la vie dans son ensemble d’un avantage essentiel, celui s’assurer la survie. Ainsi, cette constatation renforce le schéma darwinien. En effet, rappelons nous que Darwin et ses successeurs attribuent au hasard la variabilité biologique, c’est-à-dire les différences entre les individus, à l’intérieur d’une même espèce. Mais, comme ils l’ont constaté, ce hasard ne suffit pas pour expliquer qu’apparaissent de grandes différences pouvant conduire à de nouvelles espèces. Il faut associer un autre ensemble de processus, ceux de la sélection naturelle qui filtrent les individus les mieux adaptés. C’est donc le couple hasard-sélection qui explique les faits évolutifs. Mais ce n’est pas fini. En effet, on peut noter que depuis un siècle et demi un grand effort a été fait pour décrypter la mécanique de la sélection, les processus nombreux qui y participent. En revanche, peu de chose pour le hasard et ce n’est que très récemment qu’on s’intéresse à ce qui le produit, notamment ce que nous avons appelé des roulettes biologiques.
Tout cela n’est pas sans conséquences pratiques. Si la sélection est utilisée par l’homme depuis l’émergence de l’agriculture et de la domestication d’espèces animales et végétale et l’on ne peux pas nier qu’elle ait été largement bénéfique ; en revanche, le hasard, lui, a été ignoré, on a tout au plus modifié la diversification avec les marteaux-pilons que sont les facteurs mutagènes. On peut faire mieux maintenant quand nous aurons identifié et compris le fonctionnement des roulettes biologiques, très apparenté aux chaos… déterministe, et, nous pouvons l’espérer, trouver les moyens de les modifier (les accélérer les freiner, les biaiser). Par exemple, dans la gestion des écosystèmes et en particulier des zones protégées, qui sont donc laissées à un fonctionnement spontané, il faut veiller à ce que les éléments qui assurent un « bon hasard » soient protégés, si on souhaite maintenir la biodiversité de ces écosystèmes. C’est ce qu’on tente de faire de façon empirique dans les politiques de protection totale. Cependant nous savons que ces schémas qui excluent l’homme ne sont guère tenables ; il faut donc pratiquer une gestion circonstanciée et alors veiller à préserver des processus essentiels et donc ceux qui les produisent. Par exemple, le rôle des animaux disséminateurs de graines en forêt amazonienne est prouvé, il faut donc être attentif à la protection spécifique de ces animaux, sachant que tous ne le sont pas, mais peuvent intervenir de façon déterminante dans d’autres aspects du fonctionnement spontané d’un écosystème.
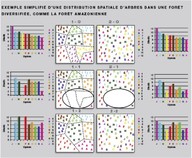
Figure. Exemple simplifié d’une distribution spatiale d’arbres dans une forêt diversifiée (on dispose d’exemples réels obtenus en forêt guyanaise). Les points correspondent à des individus de différentes espèces : chaque symbole représente l’une des six espèces présentes. Les parcelles A et B ont les mêmes diversités et abondances relatives : nombre d’espèces égal distribution par espèce identique. En 1-0, la distribution est en patchs homogènes. En 2-0, la distribution est aléatoire (comme le plus souvent en forêt tropicale naturelle). Si l’on suppose qu’une perturbation majeure touche une partie de ces parcelles et supprime les individus impactés (1-1 et 2-1), on voit immédiatement que la distribution initiale de type 1 peut conduire à la disparition locale d’espèces contrairement à la distribution 2, en supposant que la régénération se fait uniquement à partir des individus qui ont subsisté à la perturbation (pas d’immigration). On obtient alors les situations 1-2 et 2-2: 2-2 a la même diversité que 2-0 en nombre d’espèces, contrairement à 1-2 compte deux espèces de moins que 1-0 et bien sûr 2-0. Ainsi, la distribution aléatoire des individus joue un rôle d’assurance pour le maintien des espèces.
Cela étant, il ne faut pas oublier non plus que les espèces sont plus ou moins bien adaptées aux divers milieux (leurs « niches écologiques »), plusieurs d’entre elles pouvant partager un même milieu et un même espace. Ainsi, l’organisation de la biodiversité des arbres et plus généralement de la végétation se fait à plusieurs échelles : celui d’une communauté d’individus d’espèces différentes partageant un même milieu et espace (d’un hectare à quelques dizaines d’hectares), celui d’un ensemble de communautés correspondant à un écosystème (du km2 à des milliers de km2), par exemple une forêt tropicale humide, jusqu’à celui du biome qui correspond à une région bioclimatique (des milliers de km2 non nécessairement connexes). Enfin, la diversité spécifique en un lieu donné varie au cours du temps, à conditions constantes par des disparitions locales liées aux aléas de la reproduction, de la dissémination et de la germination, et si les conditions de milieux changent avec l’arrivée, mais elle aussi aléatoire, d’individus de nouvelles espèces mieux adaptés que les anciens pouvant les remplacer partiellement et même totalement. On voit donc qu’un écosystème, donc sa diversité spécifique et son organisation spatiale, évolue au cours du temps sous une double dynamique dépendant d’une part d’aléas internes (reproduction, dissémination, migration) et, d’autre part, d’aléas externes (variation des conditions de milieu), sans oublier l’apparition progressive de nouvelles espèces. En pratique, ces systèmes ne sont jamais à l’équilibre et le hasard y joue un rôle essentiel. Ce type de problème, celui de la dynamique de la biodiversité à différentes échelles de temps et d’espace, est au centre de notre action en Amazonie [7]. Le rôle joué par le hasard doit nous interpeller sur la perception scientifique qu’on a de la nature. Le terme d’écosystème est révélateur d’une vision déterministe, structurée, où le hasard n’est vu que comme un bruit de fond brouillant notre perception. Or, il joue un rôle central : l’écologie doit le reconnaître et faire alors sa “révolution copernicienne”, changer de paradigme.
Les schémas darwiniens et néodarwiniens sont aussi en train d’être complétés et j’ai pu participer récemment à ce progrès scientifique de façon incidente, presque involontaire car ce n’était pas mon objectif. Et je pense que si je l’avais voulu, je n’y serais sans doute pas arrivé, car je n’aurai pas trouvé le chemin pour y parvenir. En effet, partir de la diversité et de l’hétérogénéité des forêts intertropicales humides, s’intéresser incidemment à l’évolution, s’interroger sur le discours scientifique sur la biodiversité, tout cela sur la base d’une culture scientifique (biologique, écologique, mathématique, statistique et informatique) acquise au cours de ma vie d’enseignant et de chercheur, pour arriver à s’interroger sur le rôle du hasard, son importance dans l’évolution, mais aussi dans la dynamique des écosystèmes, et enfin tenter d’identifier les mécanismes qui le produisent, n’avait rien d’évident. D’autant plus que nous avons « traditionnellement et culturellement » une vision négative du hasard. En effet, la plupart d’entre nous ont été coulés dans le moule du déterminisme. Il a donc fallu passer au-dessus de cet écueil très idéologique [8]. C’est sans doute aussi le cas des collègues qui participent à cette nouvelle aventure scientifique [9].
Ce cheminement m’a convaincu que Théodosius Dobzansky avait parfaitement raison quand il a écrit : « Rien ne prend sens en biologie si ce n’est à la lumière de l’évolution » et me fait militer pour que l’évolution soit racontée dès le plus jeune âge et, à travers quelques expériences autour de moi, je peux témoigner que les enfants sont passionnés pour peu qu’on sache trouver les mots et les images. Cette théorie doit aussi être enseignée et de façon approfondie à tous nos étudiants des sciences de la vie. De plus, présenter simultanément l’aventure intellectuelle, mais aussi sociale, de cette théorie, de son élaboration et de sa confirmation continuelle, est aussi très stimulant et pas seulement pour contrer les dérives du créationnisme ou celles du « dessein intelligent ». Cette théorie de l’évolution est une illustration de l’intelligence des humains, mais aussi, dans certaines perversions de son interprétation, des mauvaises intentions ou même de la profondeur de la bêtise de certains. Je pense par exemple aux mouvements eugénistes ou aux excès du darwinisme social. Tout cela mérite d’être exposé à tous ceux qui nous succèderont.
Enfin, la nouvelle perception du hasard, de la structure en grande partie désordonnée des «écosystèmes», mène à s’interroger sur la vision systémique au sens d’une nature fortement organisée, d’un cosmos bien huilé avec chacun à sa place, qui prévaut encore largement. En mettant le hasard au cœur de ces «écosystèmes» ne sommes-nous pas à l’aube d’une «révolution copernicienne en écologie» ? [11]
[1] Pavé A. - Modélisation en biologie et en écologie. Aléas, Lyon, 1994, 560p.
[2] Courtillot V. and Gaudmer Y., Effects of mass extinction on biodiversity. Nature, 381,1996, 146-148.
[3] Pavé A., Hervé J.C. et Schmidt-Lainé C. – Mass extinctions, biodiversity explosions and ecological niches. Comptes rendus de l’Académie des Sciences-Biologies, 325 : 7, 2002, 755-765.
[4] Pavé A. La nécessité du hasard – Vers une théorie synthétique de la biodiversité. EDP-Sciences, 2007, 192p. En cours de traduction en anglais.
[5] Pavé A. Necessity of chance: biological roulettes and biodiversity. C. R. Biologies 330 : 3, 2007, 189-198.
[6] L’évolution. Comment les espèces s’adaptent. Les dossiers de La Recherche, 27, 2007.
[7] Voir par exemple :
-Des labos dans les arbres. Journal du CNRS. Janvier/Février 2008.
-Programme Amazonie du CNRS Phase II. Texte de cadrage et d’orientation scientifique. Septembre 2007.
[8]La citation suivante me semble bien illustrer ce propos : « Le hasard a longtemps été nié par l’Église qui y voyait une insulte aux plans de Dieu. Puis il a été nié par les savants pour qui l’univers était une mécanique bien huilée. À la limite, il y avait des lois que nous ne connaissions pas encore. » (Robert Solé, dans Le Monde littéraire du 7/12/07 à propos du livre « Qu’est-ce que le hasard ? Denis Lejeune, Ed. Max Milo, 2007).
[9] Cf, par exemple:
- Kupiec J.J. (2006). L’expression aléatoire des gènes. Pour la Science, 342, 78-83
-Mettetal J. T., van Oudenaarden A. (2007) Necessary Noise. Science, 317, 463-464.
[10] Pavé A. On the Origins ans Dynamics of Biodiversity: the Role of Chance. Springer US, 2010. 192p
[11] Pavé A. La course de la gazelle. Biologie et écologie à l’épreuve du hasard. EDP Sciences, 2011.
(rev. octobre 2011)
_______________________________________________________________________________________
L’aventure Amazonienne avec le CNRS
Cette réflexion et ce travail s’est fait en grande partie au cours de mon séjour en Guyane, d’octobre 2002, mandaté par le CNRS pour implanter un dispositif de recherche et renforcer notre station de terrain. Dès 2004, un programme interdisciplinaire était créé pour accompagner cette implantation et élargir la communauté scientifique travaillant en et sur la Guyane. Cette aventure, une véritable “success story” est en partie racontée dans un ouvrage, joliment illustré:
[12] Alain Pavé et Gaëlle Fornet. Amazonie, une aventure scientifique et humaine du CNRS. Galaade, 2010.
Actualités
 Mis à jour le 19 août 2021 - contact : [email protected]
Mis à jour le 19 août 2021 - contact : [email protected]
Site en ligne
Vous trouverez sur mon nouveau site Internet une présentation de mon parcours professionnel et quelques informations personnelles et ci-dessous mes principaux ouvrages. Des articles sont également
téléchargeables (cf. fin de la page d'accueil et la page : choix de publications à télécharger), la plupart ont été publiés dans des revues, mais quelques récents, plutôt des articles d'opinion,
sont accessibles aussi. Si vous vous en inspirez pour vos propres travaux, n'oubliez pas de les citez. Merci d'avance !













